



Introduction
This grant proposal will advocate for funding in order to study the impacts of fire on nitrogen cycling and phosphorus cycling in the chaparral ecosystem. We are asking for $10,000 to research changes in nitrogen and phosphorus content in the soil of the California chaparral over the period of two days. It will require one year to study the nitrate and ammonium levels of post-burn soil. Research would take place on two separate occasions: pre- and post-fire.
The chaparral ecosystem is found in five parts of the world with a Mediterranean forest, woodland, and scrub biome: California, Chile, western South Africa, western Australia, and the Mediterranean Basin. They are all located between 30° and 40° North and South of the Equator. Chaparral ecosystems share a Mediterranean climate, which is notable for hot, dry summers along with cool, rainy winters (Schlesinger 1985). During the spring, fall, and autumn months, the biome receives very little rainfall. In California, chaparral ecosystems exist within the California chaparral and woodlands ecoregion (Olson & Dinerstein 2002). Common plants found in this region include scrub oak, cypress trees, oak savannas, and grasses. This ecoregion has been subdivided into the California coastal sage and chaparral, the California montane chaparral and woodlands, and the California interior chaparral and woodlands (Underwood et. al 2018). While many of the plants are drought tolerant due to climate conditions of California, they are quite susceptible to natural and anthropogenic fires.
A wildfire is defined by the U.S. National Interagency Fire Center as “the unplanned ignition of a wildland fire (caused by lightning, volcanoes, or unauthorized or accidental human-caused fires) and escaped prescribed fires” (Brusentsev and Vroman 2017). Fires are becoming increasingly prevalent and intense due to warming temperatures and drying soils. Although fires are a naturally occurring process, human-induced climate change has escalated wildfire stress variables. A lack of moisture coupled with weather conditions such as wind velocity and sustained winds are good predictors of successful fire spread (Weise et. al 2005). Changes in climate extremes, such as drought, low humidity, and high temperatures can enhance the risk of fire (Narayanan et. al 2016). Due to climate change, larger fires will become more common; it will primarily impact vegetation in the western United States. Wildfires have become more extreme due to an increase in global temperatures. Westerling (2006) found that as the spring season is arriving earlier due to the transforming climate, the length of the fire season is changing. Just as well, the increase in housing development near dry vegetation has put both people and the environment at a greater risk of wildfire occurrence (Brusentsev and Vroman 2017). Human activity is responsible for 95% of wildfires in California.
Chaparral ecosystems maintain a deficit in key nutrients. The most important nutrients that limit net primary production in these ecosystems are nitrogen and phosphorus (Rundel 1983). As such, plant species found in chaparral communities often are adapted to soils that have a low amount of nitrogen and phosphorus.
Hypotheses
1: Nitrogen Cycling
Fire in Chaparral ecosystems (specifically soil, plants, and plant litter) will cause a decrease in nitrogen, but ammonium and nitrate will increase.
Nitrogen is the nutrient most likely to hinder plant growth (Barro and Conard 1991). Ash deposition and nutrient volatilization are the two main effects of fire on the nutrient cycle. When temperatures approach 200°C, volatilization begins. Despite the fact that the overall nitrogen pool shrinks with each fire, the concentration of accessible nitrogen in the form of ammonium and nitrate rises. However, as Barro and Conrad (1991) points out, the mechanics of how available nitrogen is saved the first year following a fire and how it is replaced later in the successional cycle are not well known.
In ecosystems such as deserts, grasslands, tundra, and forests, the nitrogen balance is generally not disturbed by fires (Rundel 1983). This sets the chaparral ecosystem apart, as there have been studies that indicate change in nitrogen levels after fires.
2: Phosphorus Cycling
Fire in Chaparral ecosystems will cause an increase in available phosphorus in soil, plants, and plant litter.
Compared to the nitrogen cycle, the relationship between chaparral fires and the phosphorus cycle is a less researched area. The inputs for phosphorus include “weathering, inorganic solutes in precipitation, dry deposition, and, to a lesser degree, decomposition” (Parker 2016). Available phosphorus increases after a fire, albeit to a lesser extent than nitrogen. Phosphorus, unlike nitrogen, is not lost from the chaparral ecosystem after a fire.
Rundel (1983) indicates that different studies of phosphorus in post-fire chaparral have had varied results, and he suggests that this is due to “differences in phosphate bonding or the fixing capacity of different soils, by variable immobilization of phosphorus by soil microbes, and by variable fire intensity”. Given the apparent differences in studies, it is important to perform new research in the area using modern tools, practices, and techniques. If it is necessary, we can build off previous data by analyzing past research and distinguishing between different soil types.
Approach
The field experiments will be conducted at Cuyamaca State Park in San Diego County, California. This site was chosen because (1) it has one of the largest chaparral ecosystems in California (2) there is ample space to conduct the experiment and (3) given the location’s similarity in temperatures, weather conditions, and soil type to other areas with chaparral ecosystems, the research performed in Cuyamaca State Park may be representative of the global chaparral ecosystem.
The pre- and post-fire field collection of nitrogen and phosphorus on soils, plants, and plant litter will be conducted simultaneously over the span of two days. In order to study the levels of ammonium and nitrate, we will keep track of the content levels over the course of one year. After the first round of field collection, we will prescribe a controlled burn to the given plots of land that we have determined to use. All nitrogen and phosphorus content will be measured in kilograms per hectare, while ammonium and nitrate will be measured in micrograms of nitrogen per gram of oven-dry soil. Due to differences in flammability, vegetation will be separated by (1) live twigs (2) live stems and (3) dead twigs and stems (De Bano and Conrad 1978). Distinguishing between such is important for the purpose of maintaining the accuracy of the experiment. Plant litter will be sampled through a field study technique that applies randomness over the span of a hectare. Soil will be collected at the O horizon (humus) at depths between 0 to 1 cm and 1 to 2 cm. It will be sampled at locations where plant litter exists.
In the laboratory, we will measure nitrogen, ammonium, and nitrate using the
micro-Kjeldahl technique (Moore 1938). P will be measured by (1) saturating with magnesium nitrate (2) dry ashing (3) using the vanadate-molybdate-yellow method to analyze the ashed material calorimetrically. In the soil samples, we will continue to use the micro-Kjeldahl technique to measure nitrogen. Sodium carbonate fusion will be used to measure the soil’s phosphorus.
The experiments will require the following instruments and materials: one firestarter, one pulverizer, one Micro Kjeldahl Apparatus from Labconco.com, one adjustable temperature laboratory hotplate, 10 beakers with a capacity of 250 mL, one nickel crucible with a capacity of 50 mL, 16 pipettes, 8 tweezers, one stack of pH paper, and one pair of tongs.
Expected Results
We expect that the nitrogen and phosphorus content in soils, plants, and plant litter will decrease significantly one day after the fire. We expect that the ammonium and nitrate content of soil will gradually increase and decrease over a one-year time frame. We believe that the nitrogen cycle and phosphorus cycle will change because chaparral fires affect the ecosystem’s nitrogen balance. However, it is expected that the nitrogen cycle will be most impacted by the fire. The combination of the chaparral’s susceptibility to fires and the ecosystem’s lack of nutrients makes an imperative case for environmental protections to continue and expand. Housing expansion into chaparral environments has been a proven cause of deadly wildfires. Moreover, governments around the world must take actions aimed at mitigating the climate crisis and preventing global temperatures from increasing. It would be reckless to assume that the chaparral will still exist if climatic conditions continue to change.
Tables and Figures
The tables and the two bar graphs are based on data collected by Debano and Conrad (1978). It can serve as an adequate comparison to the results of our study and it can help us to distinguish any differences in chaparral nutrients pre- and post-fire from 1978 and the present.
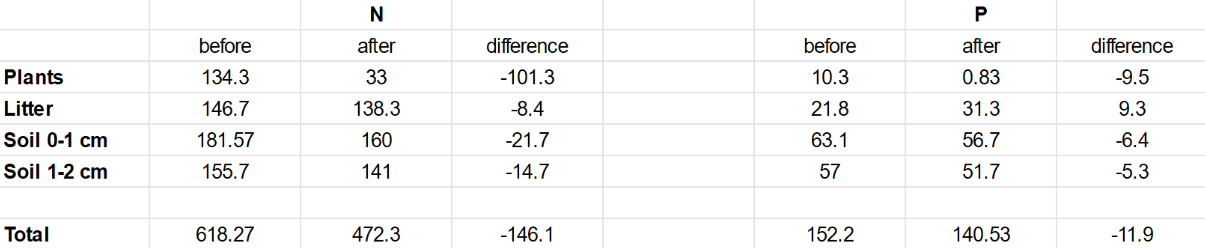
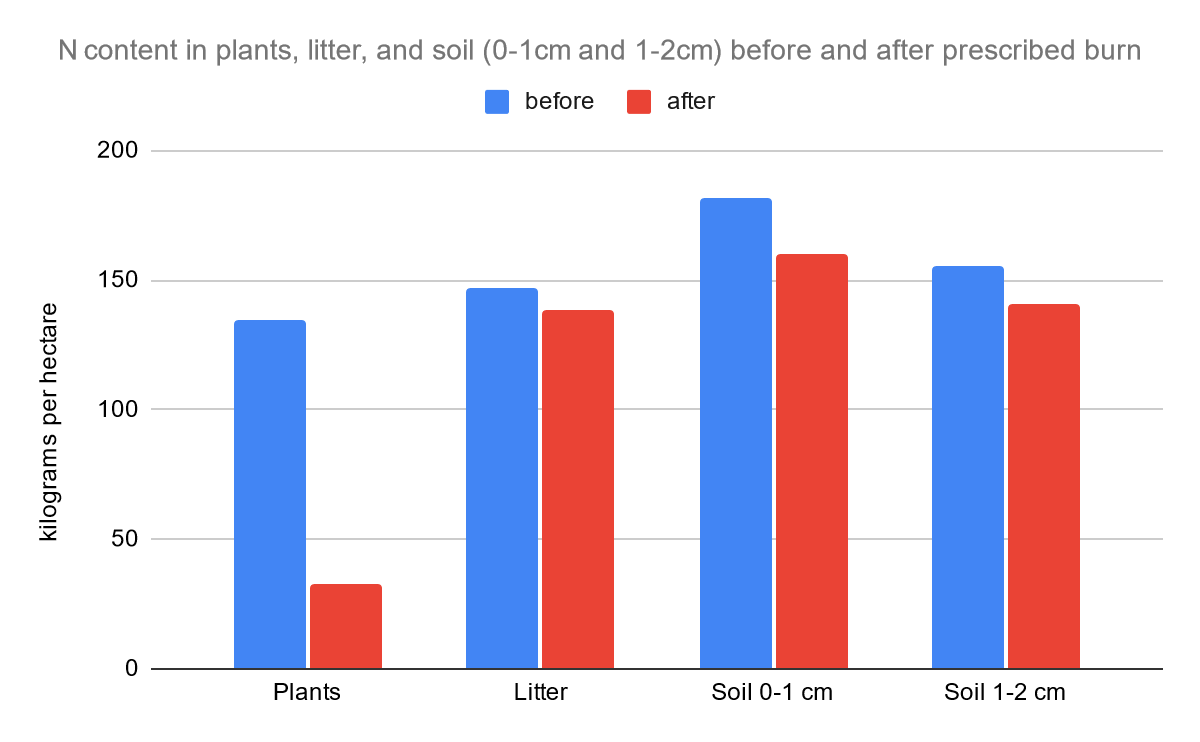
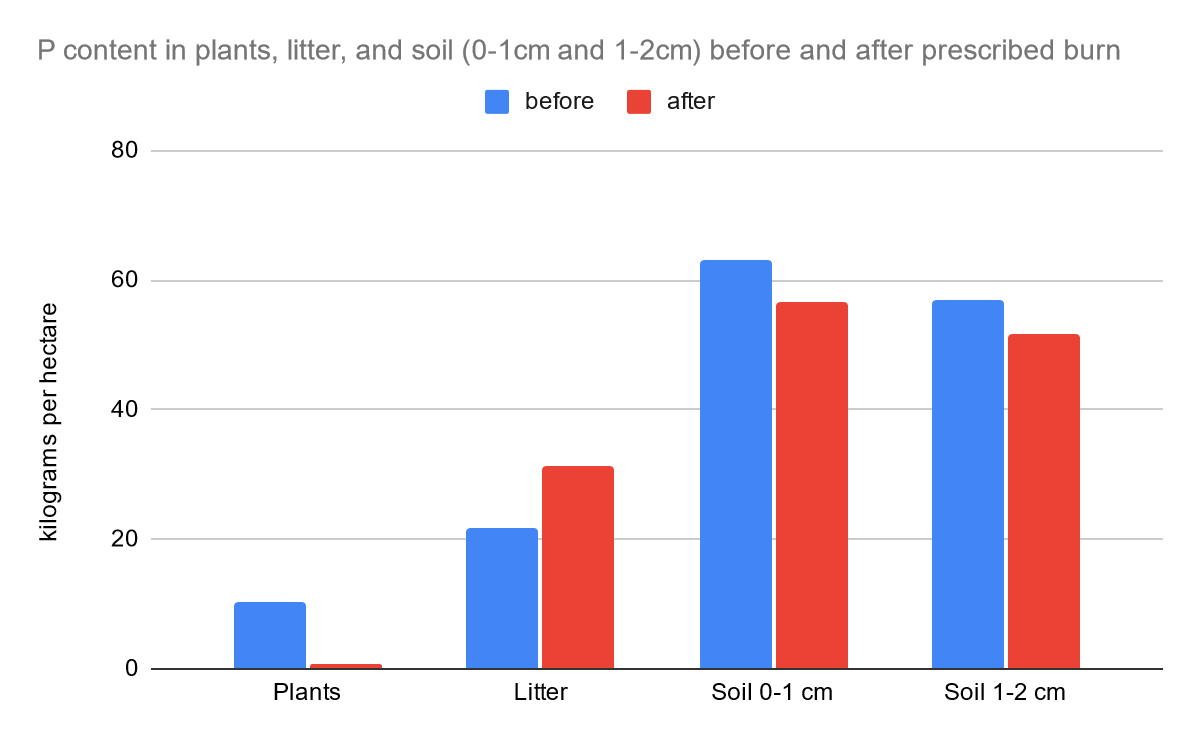
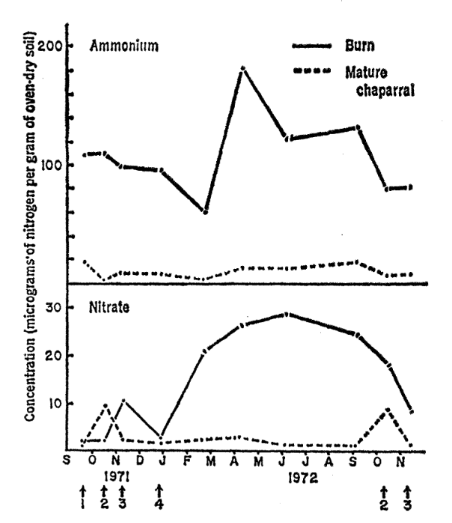
The chart above was provided by Rundel (1983). This is helpful for our research in the nitrogen cycle because nitrate and ammonium are key nutrients. It is clear that the level of ammonium and nitrate are low in chaparral ecosystems as mature adults. When the fire begins, the level of ammonium immediately increases, and the level of nitrate is bell-shaped in the increase and decrease.
References
Barro, S., & Conard, S. (1991). Fire effects on California chaparral systems: An overview. Environment International, 17(2-3), 135-149. doi:10.1016/0160-4120(91)90096-9
Brusentsev, V., & Vroman, W. (2017). Disasters in the United States: Frequency, costs, and compensation. Kalamazoo, MI: W.E. Upjohn Institute for Employment Research.
Debano, L. F., & Conrad, C. E. (1978). The Effect of Fire on Nutrients in a Chaparral Ecosystem. Ecology, 59(3), 489-497. doi:10.2307/1936579
Moore, R. H. (1938). A Micro-Kjeldahl Method Including Nitrates. Botanical Gazette, 100(1), 250-252. doi:10.1086/334778
Narayanan, A., Willis, H. H., Fischbach, J. R., Warren, D. E., Molina-Perez, E., Stelzner, C., . . . LaTourrette, T. (2016). Characterizing National Exposures to Infrastructure from Natural Disasters: Data and Methods Documentation
Olson, D. M., & Dinerstein, E. (2002). The Global 200: Priority Ecoregions for Global Conservation. Annals of the Missouri Botanical Garden, 89(2), 199. doi:10.2307/3298564
Parker, V. T. (2016). Ecosystems of California. Oakland, CA: University of California Press.
Rundel, P. W. (1983). Impact of Fire on Nutrient Cycles in Mediterranean-Type Ecosystems with Reference to Chaparral. Mediterranean-Type Ecosystems Ecological Studies, 43, 192-207. doi:10.1007/978-3-642-68935-2_11
Schlesinger, W. H. (1985). Decomposition of Chaparral Shrub Foliage. Ecology, 66(4), 1353-1359. doi:10.2307/1939188
Underwood, E. C., Franklin, J., Molinari, N. A., & Safford, H. D. (2018). Global Change and the Vulnerability of Chaparral Ecosystems. The Bulletin of the Ecological Society of America, 99(4). doi:10.1002/bes2.1460
Weise, D. R., Zhou, X., Sun, L., & Mahalingam, S. (2005). Fire spread in chaparral—go or no-go? International Journal of Wildland Fire, 14(1), 99. doi:10.1071/wf04049
Westerling, A. L. (2006). Warming and Earlier Spring Increase Western U.S. Forest Wildfire Activity. Science, 313(5789), 940-943. doi:10.1126/science.1128834